Research
We develop computational and mathematical models, as well as data analysis tools, toward a quantitative understanding of the molecular mechanisms of cell division, polarization, and motion. These fundamental cell biological processes involve interactions of the cytoskeleton with membranes, as well as the organization of signaling systems on membranes. This organization is shaped by and adapts to internally or externally generated flows and mechanical forces.
Our biophysical modeling spans both the protein and cell scales. It aims to describe the emergence of complex subcellular organization through molecular interactions. This work is carried out in collaboration with experimentalists who use techniques and model systems suited to addressing these fundamental aspects.
Research themes:
ACytokinesis
Cytokinesis is the final step in cell division when a dividing cell physically separates into two. During cytokinesis, animals and fungi assemble a contractile ring containing actin filaments and the motor protein myosin to separate into two daughter cells.
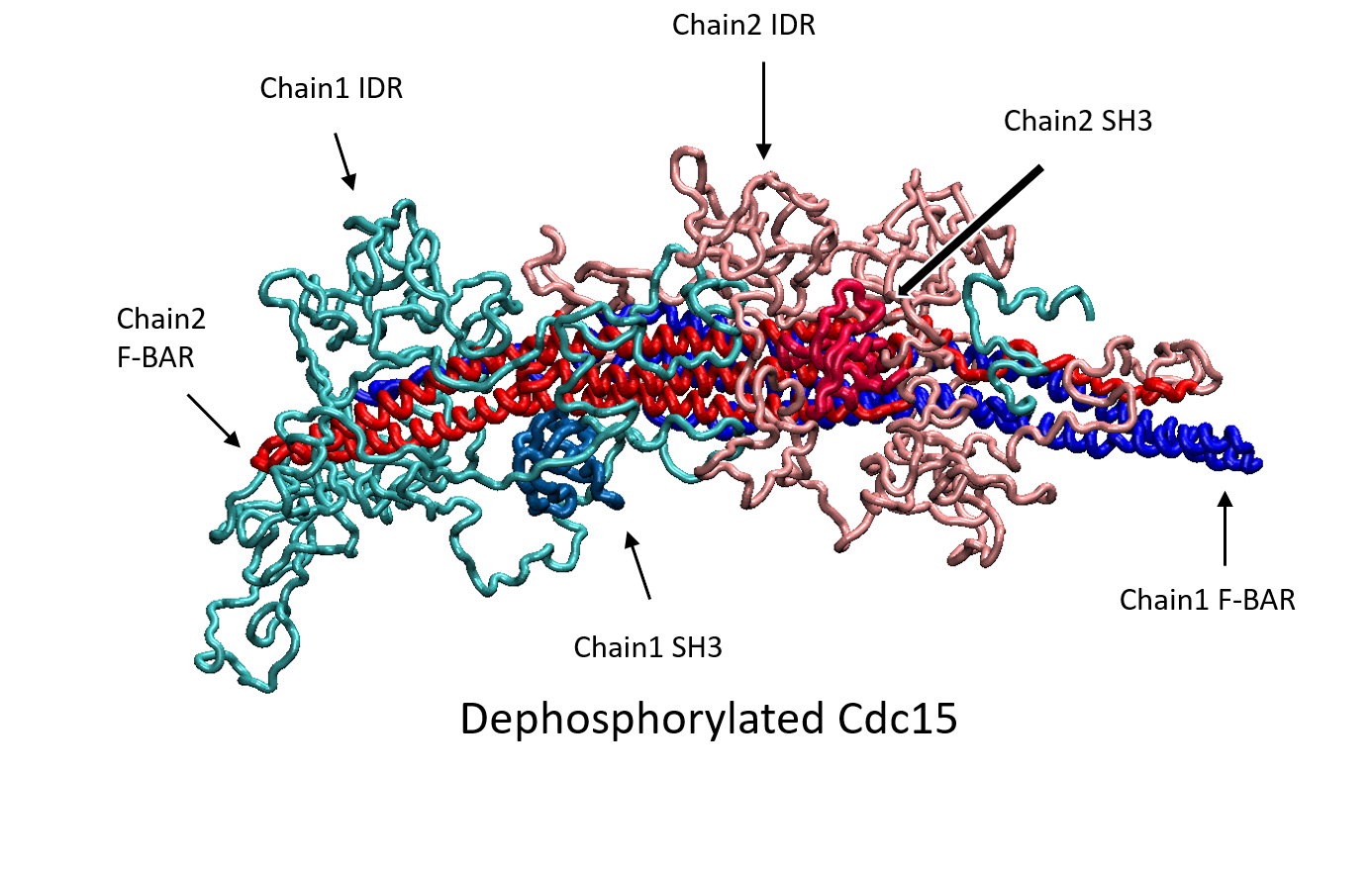
Quantitative investigations in fission yeast have revealed the role of "nodes", macromolecular complexes on the medial plasma membrane, in orchestrating cell cycle progression and cytokinesis. The nodes serve as an indicator of overall cell size during interphase and condense into the actomyosin contractile ring through a "search, capture, pull, and release" mechanism. We develop models to understand how molecular architecture and liquid-liquid phase separation aids in node organization through dephosphorylation of protein Cdc15 and other activation pathways.
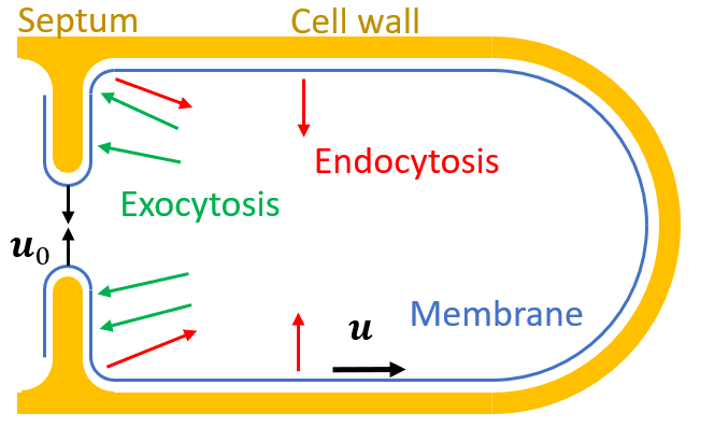
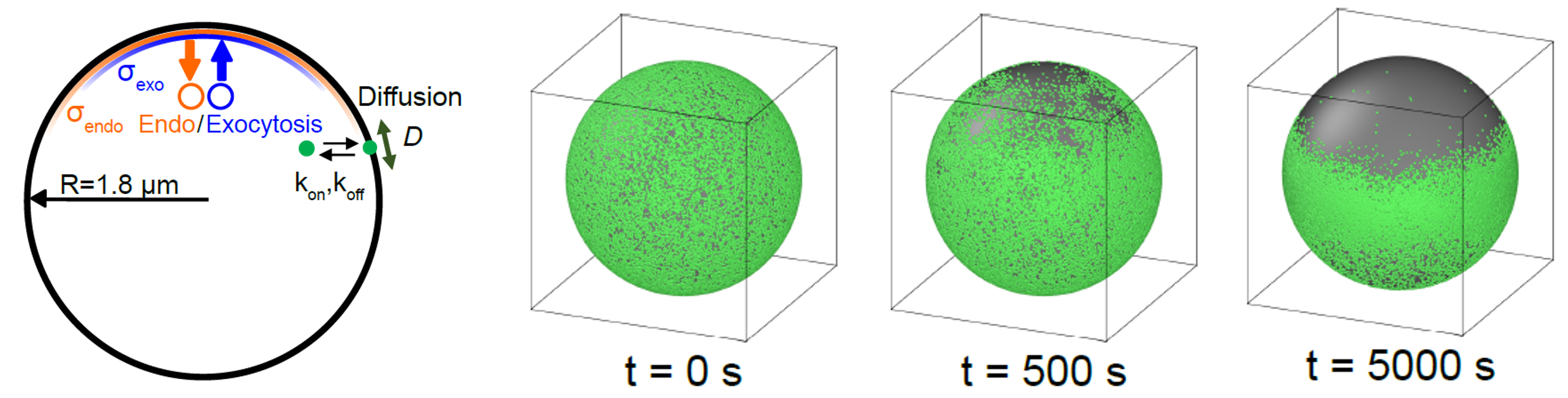
During cytokinesis, the plasma membrane must undergo two-dimensional fluid-like deformations while preserving mechanical integrity. We develop biophysical models to understand how spatial distribution of exocytosis and endocytosis regulates cytokinesis while maintaining membrane integrity.
BCell and Organelle Motility
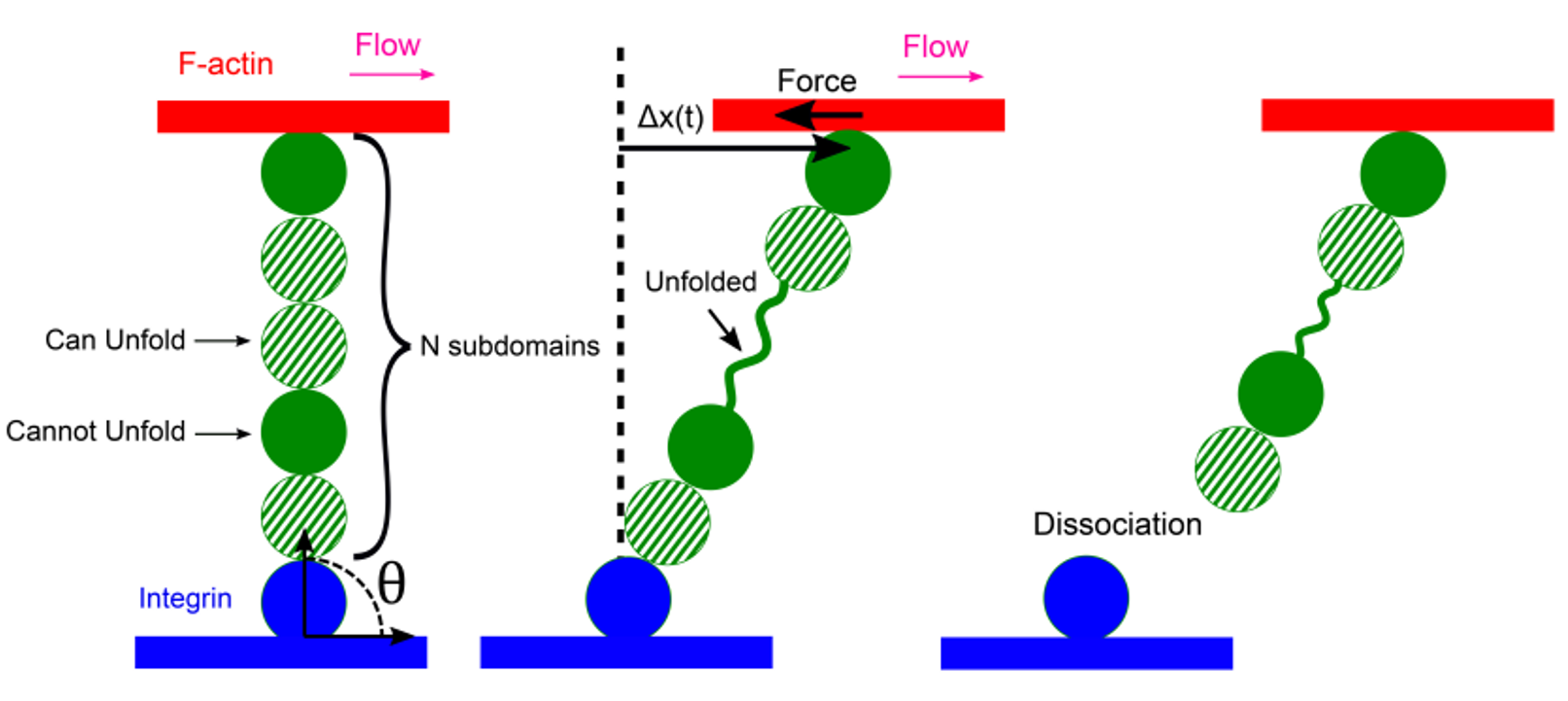
Actin filament polymerization and depolymerization provides a mechanism for cell and organelle movement and change of shape. Cellular dendritic actin networks generated through Arp2/3 complex branching off the sides of pre-existing actin filaments typically work by (a) a force generating polymerizing front, (b) distributed remodeling through severing, actin oligomer diffusion and annealing, (c) adhesion clutch and myosin pulling. We study these mechanical and kinetics processes with computational methods appropriate for the relevant lengthscales and timescales, in collaboration with the experimental group of Naoki Watanabe (Kyoto University). We have developed filament-level computational models to explore the mechanical regulation of lamellipodial actin networks undergoing retrograde flow. We work to understand how the actin cytoskeleton engages with the extracellular matrix at focal adhesions, where the connection between integrins and actin is mediated by talin, a long protein with multiple domains that unfold under force.
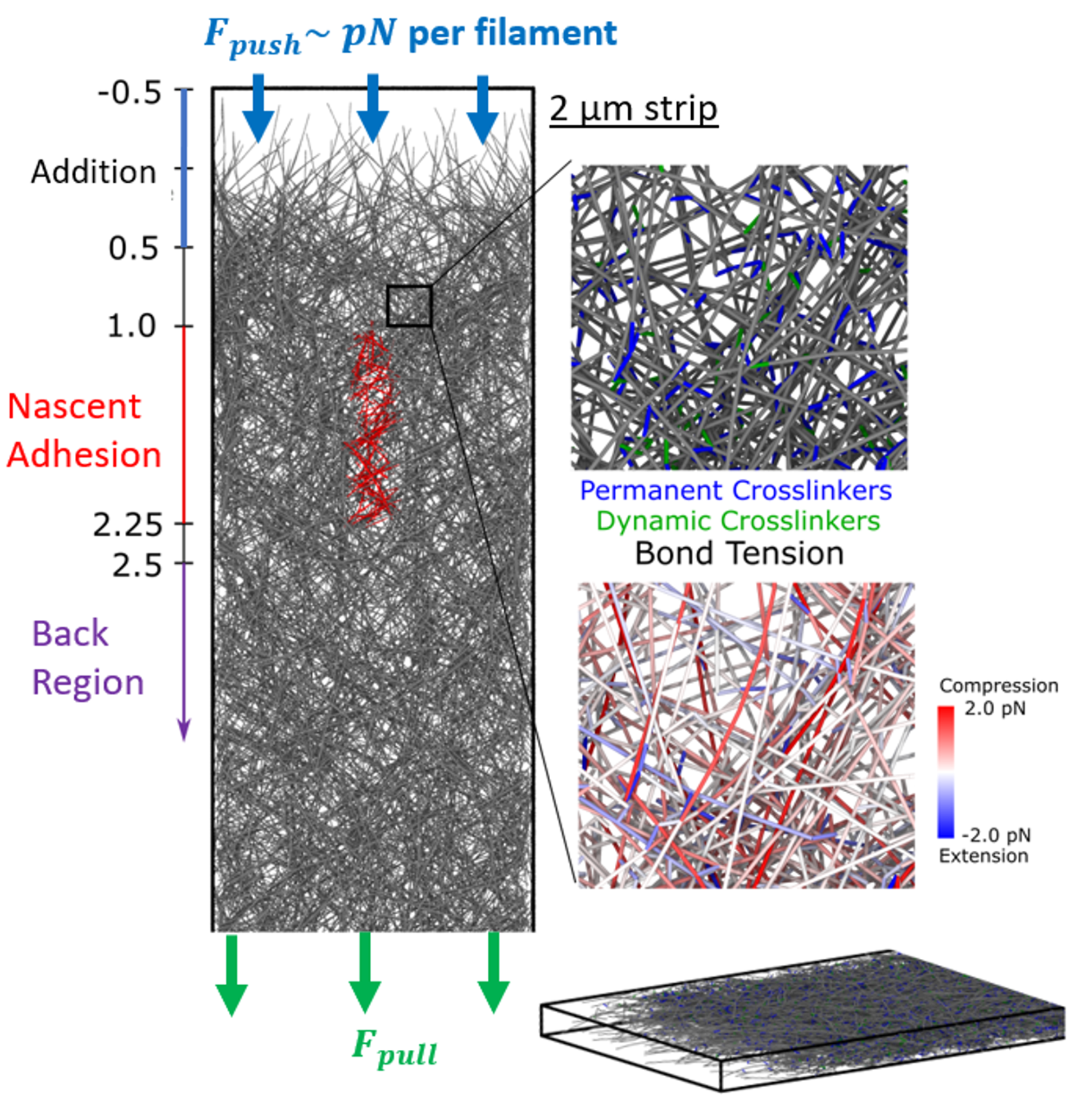
We have developed a filament-level mechanical model of Arp2/3 complex symmetry breaking and actin comet tail formation around a nucleating bead, in collaboration with the Ostap group (UPenn). The model recapitulated experimental observations, suggesting myosin-I acts as a force shaping the network’s architecture and boosting its force-generating capacity.
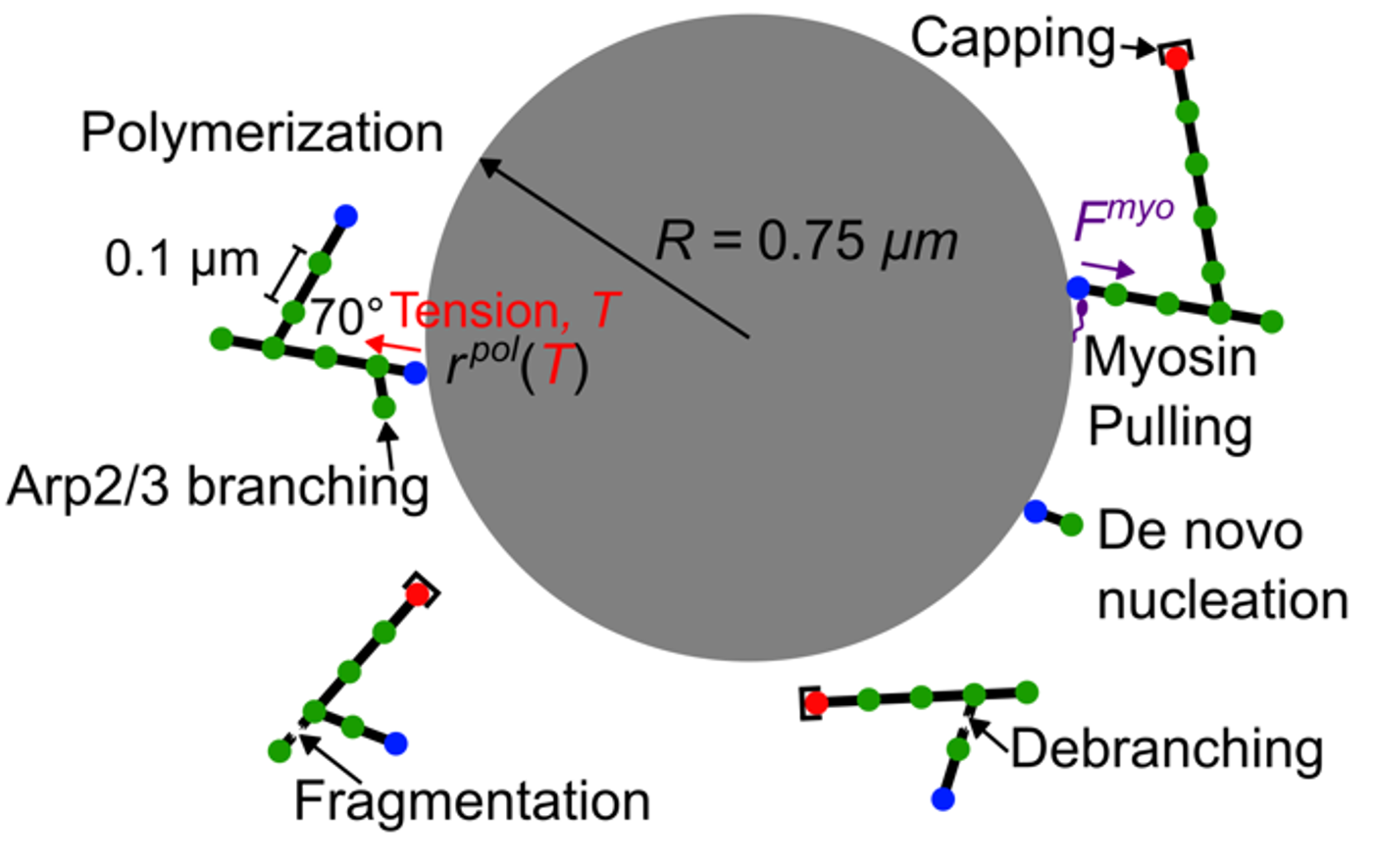
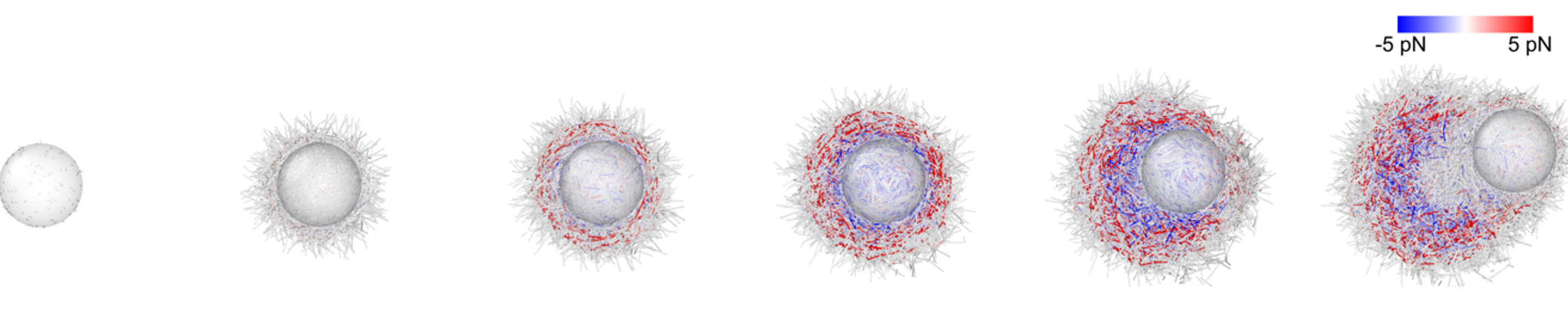
CCell Polarization
Cells establish a sense of direction for cell growth and movement using membrane proteins and the cytoskeleton. Small GTPases such as Cdc42 are key regulators of cell polarity in many eukaryotic cells, including the fission yeast, which exhibits monopolar and bipolar growth patterns. We use modeling to understand how activated Cdc42 accumulates at growth regions on the plasma membrane, with a characteristing distribution of activators (GEFs) and inhibitors (GAPs) along the cell membrane, as well as anti-correlated oscillations at cell tips. Our particle-based models of this process show how positive feedback by discretely modeled GEF activators, hydrolysis by discrete GAP inhibitors, and flow-induced displacement by exo/endocytosis are important for robust cell polarization. We also use continuum and particle-based model to understand how yeast cells polarize toward their mating partners.
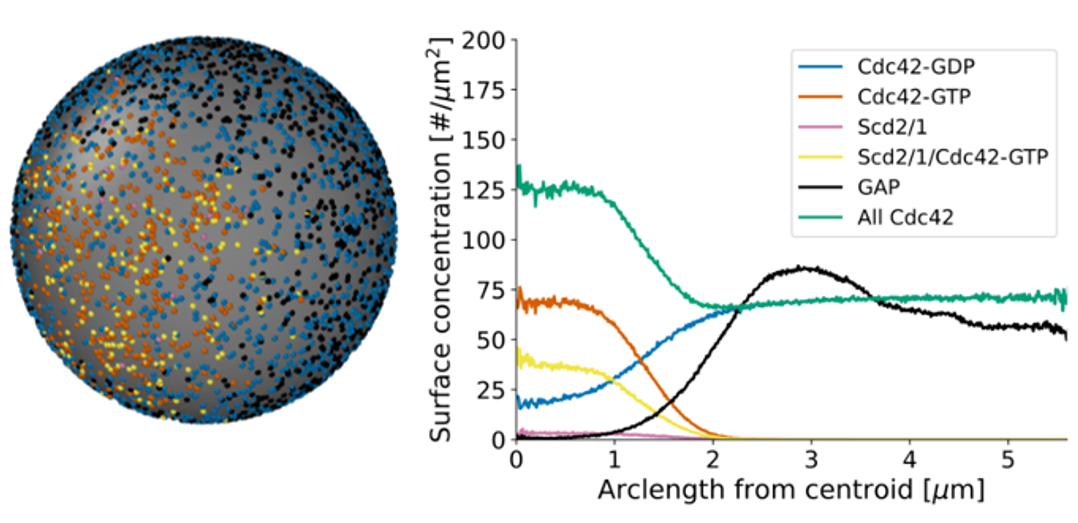
In collaborative work with the experimental group of Sophie Martin (Univ of Geneva), we have recently explored the interplay between membrane flows and activation of Cdc42. Our modeling quantifies how displacement of Cdc42 inhibitors away from cell tips via membrane flows induced by narrow exocytic vesicle delivery and wider endocytic retrieval zones is an integral part of Cdc42-driven pattern formation.
DCytoskeletal Dynamics
Our group works on the biophysical principles that govern the assembly and remodeling of cytoskeletal networks. We are interested in understanding how the free energy of hydrolysis is used by cells to power assembly and turnover of actin filament networks and bundles. We apply atomistic, coarse-grained, or chemical kinetics models to describe how actin binding proteins modulate filament dynamics and mechanical behavior. On larger length scales, we develop models describing how assemblies of filaments and motor proteins give rise to emergent structures with mechanical integrity, and their dependence on the discrete nature of crosslinker, motor, and confining interactions.
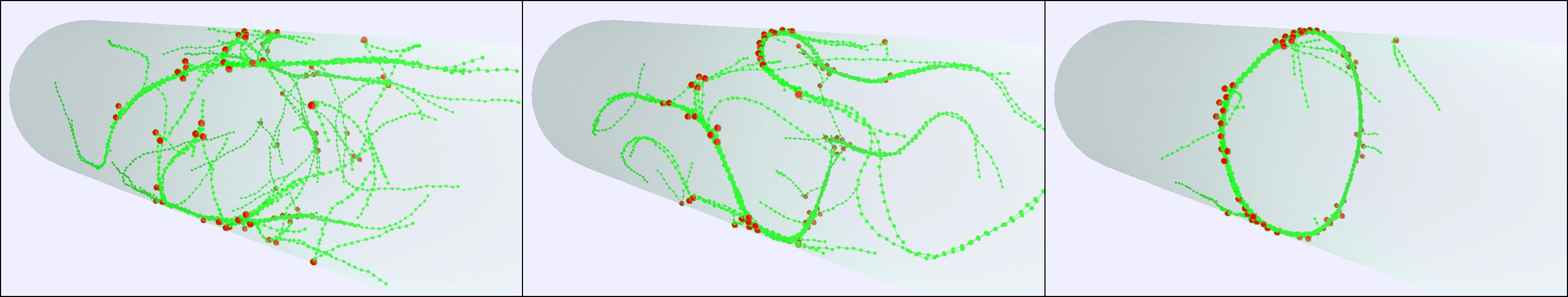
To connect our modeling with experimental image data, we have developed and maintained image analysis software such as SOAX, TSOAX and JFilament to extract and track the centerlines and junctions of filamentous networks in 2D/3D images.
We employ Brownian dynamics simulations in collaboration with the Center for Computational Biology at the Flatiron Institute using the aLENS software for large-scale actin network simulations in high performance computing environments.