The first steps in the replication process for all viruses
involve the attachment of the virus particle (the "virion") onto the
cell surface and then the entry of at least a portion of the particle (including
the genome) into the cytoplasm. The molecular interactions involved in these
steps are main factors involved in the species-specificity (narrow or wide)
of viruses.
For all viruses, the "entry problem" is to get its genome to the other side
of the cell's plasma membrane. For enveloped viruses, this almost always involves
membrane fusion between the viral envelope and a cell membrane (either the plasma
membrane or an internal vesicle membrane). For non-enveloped virions, the viral
surface protein(s) may play a role in causing some localized membrane disruption
that allows entry to occur.
We will begin our investigation of the replication processes of specific viruses
by taking a look today at the "getting in" mechanisms of several viruses. This was covered in detail in a 2004 review article in Science by Smith and Helenius titled "How Viruses Enter Animal Cells".
1. How does the enveloped influenza virus enter cells?
Enveloped viruses gain entry to cells by use of specific
viral proteins that have membrane fusion - inducing properties. The first of
these viruses for which the entry process was figured out in detail was influenza.
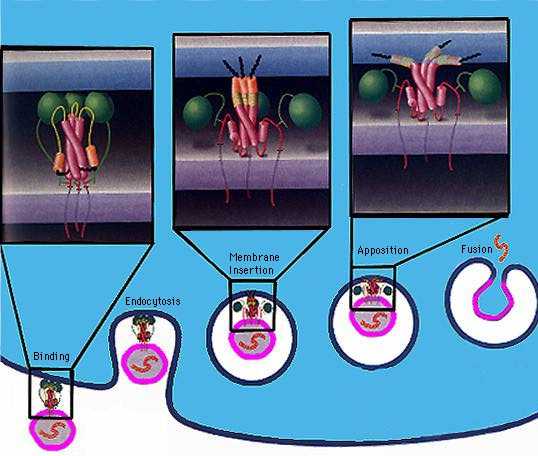
Influenza virions (see
EM pictures) bind to sialic acid (also known as N-acetyl neuraminic acid)
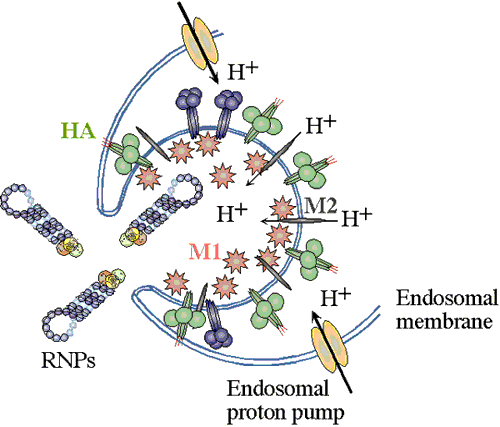
on the cell surface and are endocytosed. The endocytic vesicles become acidified (pH drops from 7 to around 5),
and at this pH, the hemaglutinin (HA) trimers in the influenza envelope undergo a structural transition. At some stage the HA monomer is cut into
two polypeptides, HA1 and HA2. When the pH drops to 5, the amino terminal end
of HA2 flips "upward" from an internal position to become exposed
to the aqueous environment. This end of HA2 (called the "fusion peptide") is
highly hydrophobic, and it interacts with the vesicle membrane and causes the
viral envelope and the vesicle membrane to fuse. (See figure.) This fusion event dumps the
viral core into the cell's cytoplasm. (See
a figure by Paul Digard,
Department of Pathology, University of Cambridge.)
A 2003 paper in Proceedings of the National Academy of Sciences by Lakadamyali
et al. concluded that the fusion event to release the viral core
actually does not happen until the endocytic vesicle has been transported the
perinuclear region of the cell. (See Figure
5 of this paper for a summary diagram.)
2. What are the details of HIV attachment
and entry?
The two viral envelope glycoproteins, gp120 and gp41 (which
are together in a complex, because they started out as a single glycoprotein,
gp160, that got cleaved) are responsible for attachment and membrane
fusion, respectively. On the cell surface, there is a glycoprotein, named CD4,
that the virus uses as the primary receptor, and one of several other cell surface
proteins is used as a "co-receptor". Many of the details of the structure and
interaction of all of these molecules were worked out in the late 1990's.
The overall sequence of events that result in HIV entry are:
* An HIV virion diffusing randomly in extracellular space
bumps onto cell.
* If this cell is displaying CD4 glycoprotein on its surface, the HIV virion
may stick, due to the specific binding of gp120 to CD4.
* A specific co-receptor protein on the cell surface interacts with the CD4-gp120
complex.
* The binding of gp120 to CD4, and the binding to co-receptor, causes a configurational
change that exposes gp41 (which had been "hidden" underneath gp120) and makes
the hydrophobic amino-terminal region of it that acts as a "fusion peptide" to be available to interact with the cell surface.
* gp41, in a "trimer of hairpins" configuration, interacts with the
cell's plasma membrane and induces fusion of the viral envelope with the plasma
membrane.
* The viral capsid enters the cellular cytoplasm as the fused membrane region
flattens out.
Research on the HIV entry process continues. The May 2006 issue of the Journal of Virology has an article by Yang et al. titled "Subunit Stoichiometry of Human Immunodeficiency Virus Type 1 Envelope Glycoprotein Trimers during Virus Entry into Host Cells".
3. How do non-enveloped virions, such as those of the icosahedral picornaviruses, adenoviruses, and papovaviruses, enter cells?
Poliovirus (and rhinoviruses, which cause about 50% of "colds") have an icosahedral structure which has been highly characterized. As we have seen, the icosahedral virion capsid consists of 60 copies each of three proteins designated VP1, VP2, and VP3, arranged in a way that forms both hexameric and pentameric vertices. The twelve pentameric vertices are composed of five copies each of VP1. A depression, or "canyon", in the structure is the region that provides the specific binding site for attachment to cells. After attachment, receptor-mediated endocytosis begins, and this leads to the RNA genome of the virus getting into the cytoplasm. The detailed mechanism of how this RNA entry step occurs is not known, but is thought to involve viral protein conformational change that opens a channel through the membrane of the endocytic vesicle. For example, there is a 2006 article by Tuthill et al. in the Journal of Virology titled "Characterization of Early Steps in the Poliovirus Infection Process: Receptor-Decorated Liposomes Induce Conversion of the Virus to Membrane-Anchored Entry-Intermediate Particles".
In the Smith and Helenius review article, Figure 2 a and c summarize our knowledge of the entry processes of adenoviruses and the papovavirus SV40.
{kind=link}
{kind=link}